|
|
Peter G. Bursztyn | |
| Fisiologia per gli sportivi |
|
|
Peter G. Bursztyn | |
| Fisiologia per gli sportivi |
1. Introduzione alla fisiologia dell'attività fisica
Introduzione
Attività fisica, collaborazione tra i vari organi
L'attività fisica: il suo miglioramento e i suoi limiti fisiologici
Il nostro corpo è una macchina straordinaria
2. I muscoli
La membrana delle cellule muscolari
Il potenziale di membrana
Il potenziale di azione
All'interno della cellula muscolare: il meccanismo di contrazione
Il meccanismo di contrazione in azione
Rigor mortis
Attivazione muscolare o stimolazione della contrazione
Controllo della forza di contrazione
Muscolatura lenta e veloce
La muscolatura a lenta contrazione favorisce il metabolismo aerobico
La muscolatura a contrazione veloce favorisce il metabolismo anaerobico
Come vengono usati i vari tipi di muscoli
Possiamo scegliere quali fibre muscolari usare o controllarle?
Tipi di muscoli e prestazioni atletiche
Doti naturali o nutrizionali: il merito è dell'ereditarietà o
dell'allenamento?
Gli effetti dell'allenamento sui muscoli
L'allenamento alla forza
Efficacia dell'allenamento alla forza
Quali risultati si ottengono con l'allenamento alla forza?
Fattori responsabili dell'aumento della forza
Modificazioni del metabolismo durante l'allenamento alla forza
Stimolazione elettrica
Allenamento alla resistenza
3. Il metabolismo
L'ATP e le fonti di energia
Ritmo di lavoro e consumo di ossigeno
Risparmio energetico giornaliero
Dove viene usata l'energia?
Come viene accumulata l'energia nell'organismo?
Dove viene immagazzinata l'energia?
Come ha inizio l'utilizzazione dell'energia?
Debito di ossigeno
La componente veloce
La componente lenta (acido lattico)
La componente lenta (eterogenea)
Sommario
Modi per accertare la buona forma fisica
Ritmo cardiaco a riposo
Ritmo cardiaco dopo lo svolgimento dell'attività fisica
Quantità massima di ossigeno introdotto (VO2 max)
Misurazione dell'inizio dell'accumulo di acido lattico nel sangue (OBLA)
Il test di Wingate per misurare la potenza sviluppata in anaerobiosi
Come migliorare le prestazioni metaboliche
Le risorse energetiche necessarie per svolgere un'attività fisica prolungata ma
moderatamente faticosa
Effetti metabolici dell'allenamento alla resistenza
Gli effetti dell'allenamento alla resistenza sul VO2 max
Il metabolismo dei grassi, la chiave della resistenza
Effetti dell'allenamento alla resistenza sulla soglia anaerobica.
Quali sono i vantaggi di chi possiede una soglia anaerobica alta?
Riepilogo
4. Fisiologia dell'apparato respiratorio
Anatomia dell'apparato respiratorio
Il meccanismo della respirazione
La difesa dei polmoni
La tensione superficiale
L'attività respiratoria
Volumi polmonari
"Efficienza" respiratoria
Rendimento respiratorio
Confronto fra il flusso di sangue attraverso i polmoni e la ventilazione
polmonare
Controllo della respirazione
Controllo chimico: anidride carbonica
Controllo chimico: ossigeno
Controllo automatico
La respirazione durante l'attività fisica
Prestazioni dell'apparato respiratorio durante l'attività fisica
La respirazione è un fattore limitante per l'attività sportiva?
Risultati dell'allenamento
Controllo della respirazione durante l'attività fisica
Riepilogo
5. Il trasporto dell'ossigeno e dell'anidride carbonica
Per il trasporto dell'ossigeno è necessaria l'emoglobina
Trasporto dell'ossigeno nel sangue
Diffusione dell'ossigeno nei muscoli
Il trasporto dell'anidride carbonica nel sangue
Il rifornimento di ossigeno è un fattore limitante per l'attività fisica?
L'allenamento alle altitudini elevate
Il "doping del sangue"
Riepilogo
6. L'apparato cardiovascolare
Parte prima: la fisiologia dell'apparato cardiovascolare
Strutture anatomiche e loro funzioni
Come scorre il sangue nei vasi
Il cuore: la sua autoeccitazione
Il cuore funziona come una pompa
La struttura dei vasi sanguigni
Tipologia dei vasi sanguigni
Le grandi arterie
Le arteriole
Sfinteri pre-capillari
I capillari
Venule e vene
Le prestazioni del cuore
La potenza del cuore
La pressione del sangue
Il controllo dell'apparato cardiovascolare
Controllo locale
Controllo centrale
Le conseguenze di un'emorragia: un esempio di controllo cardiovascolare
Parte seconda: l'apparato cardiovascolare durante l'attività fisica
La potenza cardiaca
Il ritmo cardiaco
La gettata cardiaca
La pressione del sangue
Il riflusso del sangue venoso rispecchia la potenza cardiaca
In quali direzioni agisce la potenza cardiaca?
Il controllo dell'attività dell'apparato cardiovascolare
L'efficienza dell'apparato cardiovascolare durante l'attività fisica
La velocità del flusso sanguigno durante l'attività fisica
Gli effetti dell'allenamento sull'apparato cardiovascolare
L'allenamento alla forza
L'allenamento alla resistenza
Vale la pena allenarsi?
Quali risultati si ottengono?
Il risultato globale è la somma delle sue parti
La genetica e l'apparato cardiovascolare
Riepilogo
7. Regolazione della temperatura
Premessa
Quanto calore produciamo?
La fisica del trasporto del calore
Conduzione e convezione
Irraggiamento
Evaporazione
Come regoliamo la nostra temperatura?
Che cos'è la temperatura del corpo?
Il calore ceduto dal corpo (in sintesi)
La regolazione della perdita di calore
La fisiologia della termoregolazione in dettaglio
L'irrorazione sanguigna della pelle
La regolazione del ritmo metabolico
Il meccanismo di scambio di calore controcorrente
La sudorazione
La termoregolazione comportamentale
Sommario
La termoregolazione durante l'attività fisica
La quantità di calore emesso
Come adattarsi al caldo e come allenarsi a sopportarlo
Il flusso di sangue che irrora la pelle
Il ritmo di emissione del sudore
La temperatura del corpo
Il volume del sangue
Un esempio di acclimatazione
Riepilogo
8. L'equilibrio dei liquidi durante l'attività fisica
L'attività con il caldo: tempo di cedimento
Alcune domande
Nel nostro organismo i liquidi formano comparti separati
Pressione osmotica e flusso sanguigno
Qualche esempio di come agisce l'osmosi nel nostro organismo
Durante l'attività fisica si ha una perdita sia di acqua che di sali.
Controllo della pressione osmotica
Controllo del volume
La produzione del sudore e l'equilibrio dei liquidi
La dispersione del calore e l'equilibrio dei liquidi nell'attività fisica
Si può immagazzinare l'acqua?
Produzione di sudore, equilibrio dei liquidi e prestazioni cardiovascolari
Perdita intenzionale di acqua
Prestazioni cardiovascolari, termoregolazione
e colpo di calore improvviso
Importanza del sale nella sudorazione
La sudorazione nelle persone non acclimatate: il sudore salato
Massimi di sudorazione
Da dove proviene il sudore?
La sudorazione nei soggetti acclimatati al caldo: il sale è prezioso
Il sale può essere prezioso, anche se costa poco: salare o non salare?
La reidratazione: la sete e il bere dopo la disidratazione
Le bevande per gli sportivi più comuni in commercio:
reidratazione con aggiunta di energia
Vestirsi a seconda dei climi
9. Farmaci, ormoni e altre sostanze: migliorano le prestazioni?
Premessa
I test antidoping
Eseguire i test non è semplice
I farmaci e gli ormoni più comunemente usati dagli atleti
Gli stimolanti
I diuretici
Gli analgesici e gli antiinfiammatori
Gli anabolizzanti
Gli ansiolitici
Pozioni "magiche": svariati prodotti da banco e sostanze legali
La respirazione dell'ossigeno
Il potassio e il calcio
Il cloruro di sodio (il sale da cucina)
Il bicarbonato di sodio (il lievito)
Gli aminoacidi
Le endorfine
Riepilogo
10. L'attività fisica vi mantiene in buona salute?
Che significato ha l'espressione "vi mantiene in buona
salute"?
L'allenamento alla forza
L'allenamento alla resistenza
Malattie cardiovascolari
Le malattie delle arterie coronarie e l'attività fisica
Che cosa sono le patologie delle arterie coronarie?
Che influenza ha il nostro stile di vita su ciò?
L'attività fisica e l'ipertensione
La pressione del sangue durante l'attività fisica
La pressione del sangue dopo l'attività fisica
L'attività fisica e l'osteoporosi
Il trattamento standard
Conseguenze dell'attività fisica
Gli effetti dell'attività fisica nelle donne in postmenopausa
Gli effetti di un'intensa attività fisica
Le possibili conseguenze dell'amenorrea cronica
Il caso delle danzatrici
Riassunto
L'obesità e l'attività fisica
L'attività fisica e il ritmo metabolico
L'attività fisica può far perdere peso?
Perché no?
Questo significa che l'attività fisica non serve per dimagrire?
I danni muscolari prodotti dall'attività fisica
I danni e le malattie muscolari
La contrazione concentrica ed eccentrica
I dolori, i traumi e le prestazioni
Gli effetti dell'allenamento
La guarigione
Sommario
Il colpo di calore
Quali sono le cause del colpo di calore?
La pericolosità del colpo di calore
I trattamenti
Il crampo da calore
Conclusione
Appendice
Glossario
Indice analitico
Bibliografia essenziale
PASSI SCELTI
Il trasporto dell’ossigeno e dell’anidride carbonica
Come vengono trasportati l’ossigeno e l’anidride carbonica nel sangue? L’immissione dell’ossigeno e l’espulsione dell’anidride carbonica modificano il rendimento? Il trasporto di questi gas può essere migliorato, e in che modo? Quale risultato produce l’allenamento sui sistemi di trasporto dei gas nel sangue? Perché ad altitudini elevate è impossibile compiere gli stessi sforzi che si fanno a livello del mare?
Quando si intraprende un’attività fisica il ritmo metabolico può portare il consumo di ossigeno dal valore di base (a riposo) di 250 ml al minuto fino a un valore massimo di 4000 ml al minuto (in un individuo giovane e in piena attività). Questo aumento di sedici volte della richiesta di ossigeno deve essere supportato da un aumento di cinque volte del ritmo cardiaco. Naturalmente, la quantità di ossigeno immesso nell’organismo con il flusso sanguigno deve aumentare notevolmente per equilibrare il sistema. Cambiamenti nell’efficienza dell’immissione dell’ossigeno sono strettamente collegati all’aumento del ritmo cardiaco, alla sua distribuzione nell’organismo e alla richiesta di questo gas da parte dei tessuti.
Per il trasporto dell’ossigeno è necessaria l’emoglobina
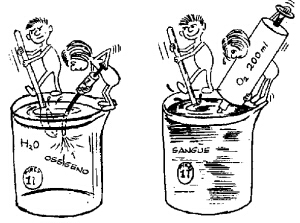
L’ossigeno non si scioglie molto velocemente in acqua. Alla temperatura del sangue arterioso, un litro di acqua esposta all’aria scioglie meno di 5 ml di ossigeno (fig. 5.1). D’altronde, a riposo il nostro organismo ha bisogno di un apporto di 250 ml di ossigeno al minuto. Se il sangue fosse costituito solamente da plasma, se ne dovrebbero pompare come minimo 50 litri al minuto per soddisfare la richiesta di ossigeno del nostro organismo, anche quando è a riposo! Dal momento che questa quantità è il doppio di quella che il nostro apparato cardiovascolare, anche con uno sforzo massimo (vedi cap. 6), è in grado di distribuire, è evidente che è necessario un sistema particolare di trasporto dell’ossigeno.
|
|
Fig. 5.1 L'ossigeno è poco solubile in acqua; infatti alla temperatura corporea soltanto lo 0,5% del suo volume si trova in soluzione. |
| Fig. 5.2 Grazie alla presenza dell'emoglobina il sangue contiene in forma solubile il 20% di tutto l'ossigeno. |
Questo particolare meccanismo è costituito da una proteina ricca di ferro, l’emoglobina, che conferisce al sangue il suo caratteristico colore rosso. Quando l’ossigeno è abbondante, l’emoglobina si combina con questo gas e lo cede quando è scarso. L’emoglobina rappresenta da oltre un miliardo di anni il migliore e più comune sistema di trasporto dell’ossigeno. Praticamente tutti i vertebrati e anche alcuni invertebrati hanno adottato questo metodo. Per mezzo di piccoli cambiamenti nella struttura della molecola, l’evoluzione ha fatto in modo che l’emoglobina si adattasse alle necessità di una grande varietà di animali di ambienti diversi, dai pesci e dai rettili, che sono animali a sangue freddo, agli animali a sangue caldo, sia terrestri che acquatici, da quelli che vivono a livello del mare agli animali adattati alle grandi altitudini.
Ogni molecola di emoglobina si unisce con quattro molecole di ossigeno, per cui ogni grammo di emoglobina si lega con 1,4 ml di ossigeno. Dal momento che ogni litro di sangue contiene 150 grammi di emoglobina, la quantità massima di ossigeno che il sangue è in grado di trasportare è poco più di 200 ml di ossigeno per ogni litro di sangue (fig. 5.2). Questa quantità è cinquanta volte maggiore della capacità del plasma di trasportare l’ossigeno. In questo modo noi abbiamo bisogno, per il trasporto dell’ossigeno, di una quantità di sangue molto minore di quanto ne occorrerebbe se nel nostro apparato cardiovascolare ci fosse solo il plasma.
L’emoglobina è una proteina di piccole dimensioni e, se si trovasse semplicemente disciolta nel plasma, i reni la filtrerebbero, facendola uscire dal nostro organismo per mezzo dell’orina. Invece l’emoglobina è contenuta all’interno dei globuli rossi (in alcune malattie, dove i globuli rossi sono danneggiati, l’emoglobina compare nelle orine, che diventano scure, come ad esempio negli stadi terminali della malaria). Inoltre, il fatto che l’emoglobina sia contenuta nei globuli rossi conferisce al sangue una viscosità minore di quella che avrebbe se l’emoglobina fosse sciolta direttamente nel plasma.
Inizialmente i globuli rossi posseggono tutti quegli organelli submicroscopici tipici della maggior parte delle cellule (nucleo, mitocondri, ecc.). Successivamente li perdono tutti, per lasciare all’emoglobina tutto lo spazio disponibile. Le cellule assumono una struttura tipicamente «biconcava», molto appiattita, in modo da disporre di una vasta superficie di diffusione (fig. 5.3). Queste cellule sono così flessibili da potersi velocemente piegare per passare attraverso i capillari: ciò si rende necessario in quanto molti capillari hanno un diametro minore di quello dei globuli rossi. Incurvandosi fortemente contro le pareti dei capillari, i globuli rossi permettono all’ossigeno di arrivare a destinazione il più velocemente possibile.
|
|
|
Fig. 5.3 L'emoglobina viene trasportata per mezzo di particolari cellule, i globuli rossi, che si infilano di stretta misura anche nei capillari più sottili. |
Trasporto dell’ossigeno nel sangue
La quantità di ossigeno trasportato dall’emoglobina dipende dalla pressione parziale del gas (la pressione esercitata da ogni gas relativamente alla pressione totale della miscela gassosa) con cui l’emoglobina si combina. La pressione atmosferica è di 760 mmHg. Poiché l’atmosfera contiene circa il 79% di azoto e il 21% di ossigeno, la pressione parziale dell’ossigeno nell’atmosfera è il 21% di 760 è cioè è di 160 mmHg. Nei polmoni, poiché l’ossigeno viene continuamente portato via, la sua pressione parziale scende a circa 100 mmHg, che corrisponde a circa il 13% della pressione totale dei gas contenuti nei polmoni. Questa è la massima concentrazione di ossigeno di tutto l’organismo. In tutti gli altri tessuti, poiché essi utilizzano l’ossigeno, la sua pressione parziale è più bassa. La pressione parziale più bassa di tutte si trova nel sangue che refluisce dai muscoli che hanno lavorato intensamente, nei quali la pressione parziale dell’ossigeno può scendere fino a 10 mmHg.
è molto importante sottolineare che il contenuto di ossigeno del sangue e la sua pressione parziale sono due cose diverse. Ad esempio, nei soggetti anemici il contenuto di emoglobina del sangue è basso, ed è anche bassa la quantità di ossigeno che può essere trasportato, anche se la pressione parziale dell’ossigeno può essere normale. Allo stesso modo, nell’avvelenamento da monossido di carbonio una parte dell’emoglobina del sangue si lega fortemente con questo gas, riducendo la quantità di emoglobina capace di trasportare l’ossigeno. Per questo motivo la persona intossicata può morire per mancanza di ossigeno, anche se i polmoni ne contengono una grande quantità totalmente disponibile1. C’è da rilevare che anche l’ossigeno disciolto nel plasma è molto scarso (figg. 5.1 e 5.4).
 |
|
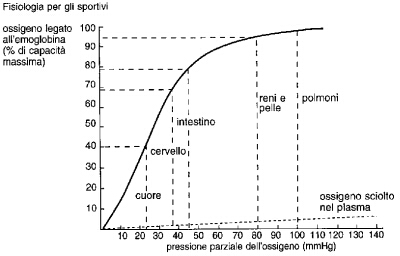
Fig. 5.4 L'emoglobina si lega con l'ossigeno in modo reversibile. Se la pressione parziale dell'ossigeno è alta (come nei polmoni) l'emoglobina si combina con l'ossigeno. Negli organi che hanno ritmi metabolici alti la pressione parziale dell'ossigeno è bassa e l'emoglobina cede loro l'ossigeno. |
Più la pressione parziale dell’ossigeno è alta, più aumenta la quantità di gas che si lega all’emoglobina, in modo da occupare tutti i punti di attacco disponibili: a questo punto si dice che la molecola dell’emoglobina è satura. Il rapporto tra la pressione parziale dell’ossigeno e l’ossigeno legato all’emoglobina è complesso (fig. 5.4). Alla pressione parziale dell’ossigeno normalmente presente nei polmoni, l’emoglobina è satura al 98%. Vale a dire che quasi tutte le molecole di emoglobina sono legate a quattro molecole di ossigeno e che in ogni litro di sangue ci sono circa 200 ml di ossigeno. Il sangue ossigenato viene trasportato dalle arterie in tutto il corpo per soddisfare le varie necessità metaboliche.
L’emoglobina cede l’ossigeno a tutte le parti del corpo che sono povere di questo gas. La quantità di ossigeno ceduto è proporzionale alla pressione parziale di ossigeno dell’organo attraverso il quale scorre il sangue (fig. 5.4). Dal momento che tutti gli organi utilizzano ossigeno, la loro pressione parziale di ossigeno è più bassa di quella del sangue arterioso. Gli organi con un metabolismo elevato tendono ad avere una bassa pressione di ossigeno. Per cui, quando il sangue arterioso scorre attraverso un organo particolarmente attivo, cede più ossigeno di quello che cede a un organo a riposo. Nella fig. 5.4 sono indicate le pressioni parziali e le corrispettive quantità di ossigeno del sangue che refluisce da vari tipi di organi.
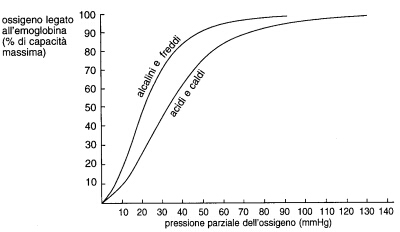
Vari fattori, oltre alla quantità di ossigeno, determinano il legame del gas con l’emoglobina. In ambiente acido, l’emoglobina cede più ossigeno (fig. 5.5). Un’alta temperatura corporea favorisce la cessione dell’ossigeno. In particolari condizioni, alcune zone del corpo con un alto ritmo metabolico tendono ad acidificarsi, per la formazione di una elevata quantità di anidride carbonica (che in soluzione si trasforma in acido carbonico). Un ritmo metabolico elevato fa anche aumentare la temperatura. Entrambi i fattori favoriscono sia il rilascio dell’ossigeno sia la sua distribuzione. La fig. 5.4 fornisce alcuni esempi di espulsione di ossigeno da parte di vari organi. Così i reni, che ricevono un notevole flusso di sangue a causa del loro ritmo metabolico, espellono soltanto l’8% dell’ossigeno di cui sono stati riforniti, mentre il cuore, che in condizioni di riposo riceve un flusso di sangue basso in conseguenza del suo ritmo metabolico, rimuove il 60% dell’ossigeno a sua disposizione.
 |
|
Fig. 5.5 La capacità dell'emoglobina di combinarsi con l'ossigeno dipende anche dalla temperatura e dall'acidità dell'ambiente. I muscoli al lavoro sono caldi e acidi e prelevano dal sangue più ossigeno del normale. Nei polmoni si verificano condizioni di temperatura bassa e di alcalinità, per cui l'emoglobina si carica di ossigeno. |
Diffusione dell’ossigeno nei muscoli
Non tutto l’ossigeno legato all’emoglobina può essere ceduto ai tessuti. L’unico sistema per permettere all’ossigeno di essere completamente liberato dal sangue sarebbe quello di far scendere a zero la pressione parziale del gas nei vari organi. Se questa situazione si verificasse, le reazioni chimiche metaboliche si bloccherebbero per mancanza di ossigeno e tutte le nostre cellule morirebbero. I nostri tessuti devono essere riforniti di ossigeno in modo da sostenere il loro ritmo metabolico e mantenere la concentrazione dell’ossigeno (o la sua pressione parziale) a un livello tale da permettere al metabolismo di svolgere il suo compito.
È difficile misurare la concentrazione dell’ossigeno contenuto nelle cellule. Sappiamo che è molto bassa nei mitocondri delle cellule muscolari che lavorano a un ritmo molto intenso. Alcuni ricercatori hanno calcolato che la pressione parziale dell’ossigeno può essere meno dell’1% del valore che questo gas raggiunge nell’atmosfera. La pressione parziale dell’ossigeno nel sangue venoso refluo è più facile da misurare e può essere utilizzata come un indicatore della diversa quantità di questo gas nei vari organi. Dalla misurazione della pressione parziale dell’ossigeno nel sangue venoso, si può constatare che i vari organi differiscono notevolmente fra di loro nella capacità di prelevare l’ossigeno dal sangue arterioso. Questo dipende da vari fattori, come ad esempio il ritmo a cui l’ossigeno viene consumato dai tessuti, la distanza tra le cellule e i capillari più vicini, la velocità del flusso sanguigno attraverso i tessuti (o la durata del contatto tra sangue e tessuto) e le caratteristiche fondamentali del tessuto rispetto al sangue (temperatura e acidità).
Il sangue che affluisce ai reni vi scorre rapidamente, poiché il compito dell’apparato escretore è quello di regolare la composizione del sangue ed espellere i prodotti di rifiuto. Di conseguenza, il sangue in uscita dall’apparato renale ha un contenuto di ossigeno molto simile a quello in entrata, con una pressione parziale di circa 60 mmHg. Il cervello utilizza solo il 30% dell’ossigeno di cui è stato rifornito, per cui la pressione parziale dell’ossigeno nel sangue refluo dal cervello è di 35 mmHg. Invece i muscoli scheletrici al lavoro possono trattenere più dell’80% dell’ossigeno ricevuto, il che corrisponde a una pressione parziale dell’ossigeno contenuto nel sangue venoso di circa 15 mmHg. La capacità dei muscoli in attività di prelevare ossigeno dal sangue è dovuta a vari fattori: alla loro alta temperatura, all’acidità, e all’abbondante apporto di sangue, che permette a ogni cellula di non essere troppo lontana da un capillare sanguigno. Tuttavia il fattore di gran lunga più importante è la peculiare perdita di ossigeno a cui i muscoli sono soggetti. Nel corso dell’attività fisica, attraverso particolari segnali inviati dai nervi vasocostrittori, tutti gli organi sono costretti ad accettare una quantità di sangue inferiore a quella di cui essi hanno bisogno, «come se l’avessero». Questo, a sua volta, fa abbassare la loro pressione parziale dell’ossigeno, costringendoli a prelevare una maggiore quantità di gas dal sangue circolante. Poiché il ritmo cardiaco durante l’attività fisica aumenta solo di cinque volte rispetto alla condizione di riposo, mentre quello metabolico si accresce di sedici volte, poter estrarre più ossigeno è di vitale importanza affinché l’apparato cardiovascolare sia in grado di sostenere adeguatamente l’attività fisica.
Un muscolo che sta lavorando può prelevare dal sangue che circola al suo interno più ossigeno di qualsiasi altro organo. Ma come può un muscolo sopportare la bassissima concentrazione di ossigeno necessaria per fare questo? Molto probabilmente le cellule muscolari sono sensibili anche alla più piccola concentrazione di ossigeno, non molto diversamente da quanto si verifica, ad esempio, nel cervello. Tuttavia, le cellule nervose cerebrali non vengono direttamente a contatto con il sangue che le irrora, perché sono circondate da uno strato protettivo di cellule gliali. D’altro canto, i muscoli sono dotati di una densa rete di capillari che permette a ogni cellula di trovarsi a stretto contatto con il flusso sanguigno. I muscoli tollerano anche assai bene l’acidità, per cui non vengono danneggiati nemmeno dall’anidride carbonica, la quale può raggiungere concentrazioni elevate durante la loro attività, consentendo anche la distribuzione dell’ossigeno. Invece il tessuto nervoso cerebrale non tollera l’acidità e ha bisogno di un ricambio sanguigno incessante per mantenere costantemente attiva la sua funzionalità.
Oltre a ciò, nei muscoli è presente una sostanza, la mioglobina, un pigmento di colore rosso. La mioglobina ha una certa somiglianza con l’emoglobina, in quanto anch’essa si lega con l’ossigeno. Il compito della mioglobina, tuttavia, non è quello di immagazzinare l’ossigeno. Piuttosto, la mioglobina aumenta la solubilità dell’ossigeno nel tessuto muscolare. Questo aumento di solubilità serve a diffondere più velocemente l’ossigeno attraverso i muscoli e ad aumentare la concentrazione dell’ossigeno nelle cellule muscolari. La concentrazione della mioglobina è molto alta nei muscoli aerobici o in quelli a lenta contrazione, nei quali sono possibili consumi molto alti di ossigeno. A causa dell’elevato contenuto di mioglobina essi assumono una colorazione scura. Al contrario, i muscoli anaerobici o quelli che si contraggono rapidamente hanno un contenuto di mioglobina molto più basso e tendono a essere pallidi. I muscoli a contrazione lenta ricevono anche un abbondante rifornimento di sangue che li rende più scuri rispetto a quelli a contrazione veloce.
 |
|
Fig. 5.6 La mioglobina si lega con l'ossigeno per mezzo di una reazione reversibile in modo simile all'emoglobina. È presente soprattutto nei muscoli a lenta contrazione (vedi cap. 2), nei quali favorisce la diffusione dell'ossigeno. |
Il trasporto dell’anidride carbonica nel sangue
A differenza dell’ossigeno, l’anidride carbonica è molto solubile in acqua. La pressione parziale di questo gas nei polmoni è meno della metà di quella dell’ossigeno e inoltre ogni litro di sangue trasporta più di mezzo litro di anidride carbonica. La quantità di anidride carbonica trasportata dal sangue è circa tre volte maggiore di quella dell’ossigeno, pur essendo questo gas trasportato dall’emoglobina.
Il trasporto dell’ossigeno da parte del sangue non provoca cambiamenti del pH (acidità). Invece, quando l’anidride carbonica si scioglie in acqua, si trasforma in acido carbonico (nel sangue questa trasformazione si verifica rapidamente a causa di un enzima che si trova nei globuli rossi). La fuoriuscita dell’anidride carbonica attraverso i polmoni diminuisce l’acidità del sangue, che invece aumenta a causa della produzione di questo gas da parte dei vari organi. Le alterazioni del pH del sangue sono da evitarsi: ci sono parecchi modi per far sì che questi squilibri siano di minima entità.
Mentre c’è un limite alla quantità di ossigeno trasportato dall’emoglobina, questo non vale per quanto riguarda la quantità di anidride carbonica che si può sciogliere nel sangue, che dipende solo dalla pressione parziale di questo gas e dalla capacità dei vari organi di sopportare l’acidità che si produce quando l’anidride carbonica si scioglie. Ad esempio i muscoli tollerano bene l’acidità, mentre il cervello non la sopporta.
Il sangue è ben tamponato contro i cambiamenti del pH. Una soluzione tampone contiene una sostanza che si combina con un acido, neutralizzandolo. Il sangue contiene diverse sostanze ad azione tampone. Una di queste è il bicarbonato di sodio (il lievito chimico dei panettieri). L’emoglobina esercita un’importante azione tampone che diventa più efficace quando cede l’ossigeno, il che si verifica proprio quando viene prodotta l’anidride carbonica. Le proteine del plasma che hanno una debole capacità di legarsi con l’anidride carbonica hanno anch’esse un’azione tampone.
Per ogni litro di sangue che passa attraverso i polmoni, vengono espulsi circa 50 ml di anidride carbonica corrispondenti esattamente al 10% di tutto il gas disciolto. Siccome la quantità totale di anidride carbonica in soluzione è piuttosto alta, mentre invece la percentuale che passa nel sangue o che esce dai polmoni è bassa, questo contribuisce a far sì che nel sangue il pH subisca solo piccole variazioni. Bisogna tenere ben presente che il controllo della quantità di anidride carbonica nel sangue arterioso è il più importante sistema di regolazione del pH a breve termine di cui dispone il nostro organismo. Nel cap. 4 si può trovare un esempio di cosa avviene nel nostro organismo quando lavora in emergenza (cioè con alti ritmi di lavoro anaerobico).
 |
|
Fig. 5.7 L'anidride carbonica è molto solubile sia nell'acqua che nel sangue. Il sangue può sciogliere un volume di anidride carbonica pari a metà del suo volume (confrontare con la fig.5.2). |
Il rifornimento di ossigeno è un fattore limitante per l’attività fisica?
La capacità dell’apparato cardiovascolare di rifornire l’organismo di ossigeno probabilmente limita la quantità di lavoro aerobico che può essere svolto. Ad esempio, ad altitudini elevate, la disponibilità di ossigeno è ridotta. Se un particolare tipo di lavoro viene compiuto a livello del mare e poi eseguito a un’altitudine elevata, lo sforzo del cuore è minore quando si effettua la prova a livello del mare, in quanto per trasportare una sufficiente quantità di ossigeno in grado di soddisfare le richieste dell’organismo occorre una minore quantità di sangue. Durante uno sforzo di notevole entità a un’altitudine elevata, il consumo massimo di ossigeno e la quantità di lavoro svolto sono notevolmente più bassi rispetto ai valori riscontrati a livello del mare. Questi valori più bassi sono da mettere in relazione alla quantità di ossigeno contenuto nel sangue arterioso, e indica che l’apparato cardiovascolare non dispone di riserve che gli permettono di compensare la carenza di ossigeno che si verifica ad altitudini elevate.
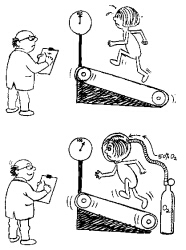
La quantità massima di ossigeno che si consuma correndo su una pedana mobile inclinata può essere aumentata del 10-12% se i soggetti respirano una miscela di gas contenente il 50% di ossigeno e l’altro 50% di azoto, invece dell’aria (che contiene il 21% di ossigeno). La frequenza del ritmo cardiaco è simile in entrambi i casi. Poiché l’ossigeno consumato e il lavoro svolto sono correlati con la capacità di trasporto dell’ossigeno, che aumenta di poco, questo fa pensare che il metabolismo aerobico sia veramente condizionato dall’apporto di ossigeno.
Sono stati effettuati degli studi su soggetti che pedalavano su di una cyclette prima con la gamba destra e poi con entrambe le gambe. Durante questi due test venne misurato il consumo di ossigeno della gamba destra. Quando anche la gamba sinistra venne fatta lavorare, il ritmo cardiaco aumentò, ma non del doppio. La quantità di ossigeno di cui la gamba destra aveva bisogno si ridusse, facendo anche diminuire il ritmo della sua utilizzazione.
 |
|
Fig. 5.8 Respirare l'ossigeno fa aumentare leggermente le prestazioni, ma a scapito della mobilità (vedi anche cap. 9, pp. 239-240). |
Per cui appare del tutto evidente che l’utilizzazione dell’ossigeno e il ritmo del lavoro aerobico sono in stretta correlazione con l’apporto di ossigeno ai muscoli. Questo fatto rende ben evidente che l’apparato cardio-vascolare rappresenta il fattore limitante nelle prestazioni di tipo aerobico.
Il sistema di distribuzione dell’ossigeno ha importanza nell’allenamento alla resistenza. Questo argomento sarà trattato nel sesto capitolo.
L’allenamento alle altitudini elevate
Dieci-quindici anni fa, un gruppo di atleti provenienti da diversi villaggi del Kenia, della Tanzania e dell’Etiopia situati ad altitudini elevate risultarono vincitori in gare di resistenza alle Olimpiadi di Città del Messico. Poiché Città del Messico si trova a 2000 metri sul livello del mare, per un certo periodo divenne consuetudine allenare gli atleti alle gare di resistenza in località poste ad altitudini elevate.
Una delle conseguenze dell’ipossia da altitudine è la formazione nei muscoli di nuovi capillari, che permettono al sangue di diffondersi maggiormente riducendo la distanza fra le zone di diffusione. Si verifica anche un aumento degli enzimi responsabili delle reazioni del metabolismo aerobico, con il risultato di aumentare la cattura dell’ossigeno. Infine, l’ipossia da altitudine stimola la produzione dei globuli rossi, facendone aumentare il numero e arricchendo il sangue di emoglobina in poche settimane. Questo accresce la capacità di trasporto dell’ossigeno, ma produce anche un aumento della densità del sangue, con il risultato di ridurre la frequenza del ritmo cardiaco. Talvolta gli indigeni delle terre situate ad altitudini elevate sono affetti da una malattia (il morbo di Monge) per effetto della quale i globuli rossi costituiscono oltre il 75% del volume totale del sangue (la norma è il 45%), sicché la viscosità del sangue aumenta di due volte.
Parecchie settimane di allenamento ad altitudini elevate possono rendere competitivi nello svolgimento di incontri sportivi gli atleti che vivono a basse quote nei confronti degli atleti di zone di alta montagna. Tuttavia, non è del tutto provato che un periodo di allenamento a grandi altezze possa costituire un vantaggio per un atleta che debba poi gareggiare a livello del mare. Probabilmente questo si verifica perché sia il consumo di ossigeno che la quantità di lavoro svolto alle altitudini elevate sono più bassi del normale. Dal momento che ad altitudini elevate non si può programmare un alto livello di allenamento, è possibile che ciò produca un leggero effetto di «deprivazione», con il risultato di contrastare ogni aumento della capacità di trasportare l’ossigeno.
Un’altra spiegazione potrebbe essere che l’allenamento in alta quota riduce l’effetto tampone prodotto nel sangue dal bicarbonato. Questo, a sua volta, può rendere gli atleti meno capaci di contrastare le elevate concentrazioni di acido lattico che si producono durante le competizioni sportive. Tuttavia, il normale contenuto di bicarbonato del sangue e il suo effetto tampone vengono di solito ripristinati in 24 ore, per cui questa spiegazione non sembra essere corretta.
In ultimo, la capacità di resistenza viene spesso limitata da due fattori concomitanti, e cioè il ridotto volume di sangue e l’aumento della sua viscosità quando l’organismo tende a disidratarsi per un’eccessiva sudorazione (vedi cap. 8). L’allenamento ad altitudini elevate aumenta la concentrazione dei globuli rossi nel sangue, per cui la viscosità del sangue degli atleti che si allenano ad alte quote tende a salire e aumenta sempre di più con l’aumentare della sudorazione.
Il «doping del sangue»
Se l’approvvigionamento di ossigeno è un fattore limitante dell’attività fisica, è evidente che l’aumento della capacità dell’apparato cardiovascolare di distribuire l’ossigeno dovrebbe migliorare le prestazioni atletiche. Gli atleti che hanno ottenuto un buon livello di allenamento nelle attività di resistenza hanno conseguito questo risultato sia attraverso l’aumento della densità dei capillari sia con un incremento dell’intensità del ritmo cardiaco. Tuttavia l’allenamento alla resistenza produce un aumento maggiore della quantità di plasma rispetto alla quantità di globuli rossi; ne consegue che il sangue degli atleti allenati ad esercizi di resistenza è un po’ meno denso, avendo una concentrazione di globuli rossi più bassa del normale.
Alcuni atleti, che hanno fatto ogni sforzo per riuscire a conseguire il massimo rendimento, si sono accorti che il loro sangue diventava meno denso e hanno cercato di aumentare la capacità del sangue stesso di trasportare l’ossigeno. Per ottenere questo risultato gli atleti praticano una tecnica che consiste nel «doping del sangue», con la quale l’atleta preleva e immagazzina all’incirca un litro di sangue almeno un mese prima di una gara importante. Il sangue che è stato tolto, così come la maggior parte delle cellule in esso contenute, vengono riformati dall’organismo nel periodo di tempo che precede la competizione. I globuli rossi immagazzinati (di norma i globuli rossi vivono 90 giorni, anche se questo dipende dalle modalità di conservazione, che può distruggerne alcuni) vengono reintrodotti nell’organismo il giorno della gara, facendo notevolmente aumentare la capacità del sangue di trasportare l’ossigeno. I globuli rossi aggiunti sopravvivono nell’apparato circolatorio per parecchie settimane: gli effetti che si ottengono con questo metodo dovrebbero durare per una o due settimane.
La reintroduzione di 400 ml di globuli rossi aumenta la capacità di trasporto dell’ossigeno del 16% circa. Tuttavia l’incremento dell’utilizzazione dell’ossigeno è variabile e può anche non verificarsi. Diversi ricercatori hanno calcolato aumenti variabili dal 5 al 15%. Poiché i globuli rossi contribuiscono a far aumentare la densità del sangue, il doping produce un aumento della sua viscosità, riducendo sia la frequenza del battito del cuore che la sua potenza. Non è facile determinare quali conseguenze si possono produrre in ogni soggetto, se cioè si verifichi un aumento della quantità di ossigeno trasportato o una minore efficienza dell’attività cardiaca.
 |
|
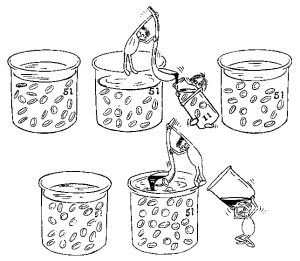
Fig. 5.9 Il sangue contiene il 45% di globuli rossi. Nel "doping del sangue" viene prelevato un litro di sangue da cui vengono sottratti, per essere immagazzinati, 450 ml di globuli rossi. Il sangue rimasto avrà un volume ridotto, ma una normale concentrazione cellulare. In un giorno o due il sangue ritornerà al volume iniziale, ma la concentrazione dei globuli rossi sarà del 36% (una debole anemia). Dopo un mese sia la quantità di sangue sia la concentrazione dei globuli rossi saranno ritornate normali. Nel sangue prelevato e conservato alcuni globuli rossi si disgregano, per cui verranno reintrodotti 400 ml di globuli rossi. Il volume totale del sangue è ora di 5,4 litri e la concentrazione dei globuli rossi è del 49%. La capacità di trasporto dell'ossigeno è aumentata e ha una durata di 2-3 settimane. |
Perciò i metodi attualmente utilizzati per il «doping del sangue» danno risultati controversi. In alcuni soggetti non si determina alcun miglioramento nelle prestazioni atletiche, mentre in altri si verifica un aumento sia della resistenza che della velocità. In campo atletico, il vantaggio anche soltanto di un centesimo di secondo può costituire la differenza fra il primo e il quarto posto, il che rende l’atleta molto interessato a utilizzare qualsiasi tecnica che gli permetta di conseguire un miglioramento anche piccolissimo. Oltre a ciò, nessun tipo di ricerca indica che il doping del sangue potrebbe peggiorare il rendimento atletico. Sembra che questa tecnica possa essere «una scommessa a senso unico». Dal momento che è difficilmente individuabile, viene ampiamente usata, pur essendo vietata.
Da un punto di vista puramente teorico, il doping del sangue può essere vantaggioso nelle gare su percorsi di media lunghezza. L’aumento della viscosità del sangue causato dalla sudorazione diventa notevole dopo un’ora di intensa attività (vedi cap. 8) e se la viscosità ha già subito un aumento a causa del doping del sangue, il rendimento atletico potrebbe probabilmente subire un peggioramento in attività sportive come la maratona o il Tour de France.
I vantaggi del doping del sangue, sempre che ci siano, potrebbero non essere dovuti a un miglioramento del trasporto dell’ossigeno. Piuttosto potrebbero essere attribuiti al periodo di tempo prima della reintroduzione, durante il quale l’atleta si è allenato con un sangue che ha una capacità di trasporto dell’ossigeno più bassa del normale: questa situazione potrebbe essere simile a quella che si verifica alle altitudini elevate, dove la disponibilità di ossigeno è ridotta.
Un’altra spiegazione degli effetti del doping del sangue è di tipo psicologico. Infatti, un atleta che è convinto di poter superare i suoi avversari per mezzo di questa tecnica, acquisisce una maggiore sicurezza e quindi ottiene un miglior rendimento.
Riepilogo
In sintesi, è probabile che la capacità dell’apparato cardiovascolare di distribuire l’ossigeno possa limitare il rendimento atletico. La reintroduzione nel sangue di un atleta dei suoi stessi globuli rossi aumenta sempre la capacità del sangue di trasportare l’ossigeno. Ciò si traduce spesso in un aumento della quantità massima di ossigeno che può essere assunto, il che permette all’atleta di migliorare le sue prestazioni per quanto riguarda la velocità, o la resistenza, o entrambe. Anche l’allenamento alle altitudini elevate produce un aumento del numero dei globuli rossi e della capacità del sangue di trasportare l’ossigeno. Questo fa migliorare il rendimento atletico nelle zone ad alta quota, ma non lo modifica alle basse quote. Tuttavia l’allenamento alle altitudini elevate raramente fa migliorare il rendimento anche alle basse quote, in quanto produce un risultato meno brillante di quello che si ottiene con il doping del sangue.
Il doping del sangue è vietato. è anche una tecnica antisportiva, poiché i suoi vantaggi (opinabili) possono essere ottenuti soltanto da quegli atleti che hanno i mezzi o la fortuna di trovare un medico poco scrupoloso. Questa tecnica presenta dei rischi se non vengono utilizzate delle particolari norme igieniche per prelevare il sangue e immagazzinarlo per lunghi periodi e per separare i globuli rossi dal sangue. Appare naturalmente superfluo far presente che gli atleti che hanno la possibilità di disporre di questa tecnica dovrebbero evitare di approfittarne se vogliono gareggiare lealmente: evidentemente, al giorno d’oggi, ruota nel mondo dello sport un grosso giro di soldi e un eccessivo desiderio di primeggiare (vedi cap. 9).
![]()
Tutti i diritti sono riservati. È vietata la riproduzione del testo in qualsiasi forma senza permesso dell’editore, salvo nel caso di citazioni o di recensioni, purché quanto in esse riportato sia conforme all’originale e se ne citi la fonte.
|
|
Via Malta, 36/8 - 10141 Torino Tel. 011-3821049 - Fax 011-3821196 |